グルコース Glucose
概要
グルコース glucose は以下の構造をもつアルドヘキソース aldohexose である(6)。D-グルコースはブドウ糖またはデキストロースとも呼ばれ,D-フルクトースとともに最も分布が広い単糖 monosaccharide である。
炭水化物 carbohydrate,糖 sugar,アルドース aldose,ケトース ketose などの言葉の意味がよくわからない場合は,炭水化物の概要のページをまずご覧下さい。
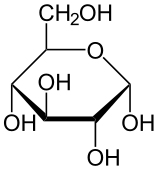
分子式 C6H12O6
分子量 180.16
構造式の書き方(α-グルコース)
- O を右上においた六員環を書く。
- 左上に CH2OH がつく。
- CH2OH の真下のみ上側に OH がつく。
- 他の部分は下側に OH がつく。
Howarth projection と呼ばれるこの書き方において,α は不斉炭素原子 C1 (リング中の O の右側の炭素) と CH2OH が逆側についているという意味である。同じ側についていれば β になる。
生物学的に重要な性質
- ヒト,マウスなど哺乳類の第一のエネルギー源である。とくに脳 brain と赤血球において重要。
- 脳が通常状態でエネルギー源にできる唯一の物質である(6)。
- 赤血球はいかなるときもグルコースしかエネルギー源にできない(6)。
- 血中で還元性を示し,タンパク質を糖化するため原則として有害である。しかし他の単糖よりもタンパク質を糖化しにくく,これが生物がグルコースを主要な糖として選択した理由の一つとされる(6)。
化学的な性質
- α 型(アルコールからの結晶)の融点は 146℃,1 分子の結晶水をもつ α 型の融点は 83 ℃(6)。
- β 型(ピリジン pyridine から再結晶)の融点は 148 - 155 ℃。
構造

直鎖状のグルコースは -(C=O)H というアルデヒド基をもつアルデヒド aldehyde の一種である。IUPAC 名で呼ぶならば 2,3,4,5,6-pentahydroxyhexanal である。Pyran は O を含む六員環である。
アルデヒドの特徴的な反応として,アルコールと反応してヘミアセタール hemiacetal を作る ことが挙げられる(6)。
R1-(C=O)H + R2-OH -> R1-(CH-OH)-OR2
ちょっと書き方が苦しいが,右辺では -R1, -H, -OH, -OR2 が C に直接結合している。なお,アセタール acetal とは R-C(OR)(OR)R という構造をもつエーテルの一種で,アルデヒドまたはケトンにアルコールを縮合させると得られる。
グルコースは分子内に -OH 基をもつので,この反応が分子内で起こって環状化する。水溶液中では α-glucose(正確には α-D glucopyranose),D-glucose(直鎖状),β-glucose(β-D glucopyranose)が平衡状態を保っている。
それぞれの分子種の割合は,NMR で調べることができる。-(C=O)H の水素原子に特徴的なピークが H- および13C-NMR のいずれでも観察されないことから,直鎖状のグルコースは極めて少ないことがわかる(17)。一般に,平衡状態では α-アノマーが 36%,β-アノマーが 64% を占めると言われている(17)。
代謝
解糖 Glycolysis
グルコースは多くの生物で主要なエネルギー源であり,これを分解して ATP を生み出す経路,解糖系 glycolysis の経路は広く保存された代謝経路である。
"Glycolysis is the sequence of reactions that metabolized one molecule of glucose to wto molecules of pyruvate with the concomitant production of two molecules of ATP" (6).
- グルコースが 2 分子の ATP を生み出しつつ各種酵素で分解され,ピルビン酸に至るまでの反応(6)。
- 解糖が酸素を必要としないのは,大気中の酸素濃度が増える前に生まれた経路だからと考えられる(6)。
- 解糖系を逆に辿ると糖新生経路に近いが,いくつかの高エネルギー反応の部分で迂回経路を通る(6)。
- 解糖と糖新生は,同じ細胞内で同時には起こらない(6)。
詳細は 解糖系のページ を参照のこと。
組織ごとのグルコース代謝の特徴
グルコースは哺乳類の脳の主要なエネルギー源である。解糖や TCA 回路など基本的な代謝経路は他の臓器と共通する部分が多いので,下記の筋肉のグルコース代謝の図が参考になる。詳細はこちら。
脳は代謝が活発な器官で,その重さから予測される量の約 10 倍早く代謝を行っている(13I)。
> 脳内に Glc は 1 mmol/kg wet weight 程度存在し,血液への注射で 9 mmol/kg 程度まで増える(10)。
> 0.25-0.50 µmol/min/g の速度で酸化されている(1)。酸化の60-90%は glutamatergic neuron で起こる。
> 老化でニューロンの Glc 代謝 (TCA cycle flux) は約30%低下するが,gliaのそれは同程度増大する(5)。
血糖値の維持
哺乳類(文献6, 9)
グルコースは哺乳類の重要なエネルギー源であり,とくに赤血球と脳 brain はエネルギーの大部分をグルコースに依存している。そのため,血中グルコース量は低下し過ぎないように調節される必要がある。
また,グルコースはタンパク質をカルボニル化して機能を低下させるため,必要以上の量が血中に存在するのも好ましくない。したがって,血糖値は狭い範囲に保たれる必要がある。
この調節は,肝臓および腎臓での糖新生 gluconeogenesis と,筋肉や脂肪組織などの糖取り込みによって行われている。インスリン insulin やグルカゴン glucagon が重要な制御ホルモンである。
正常な血糖値の範囲
血糖値は食後に急増するので,絶食時の値で比較するのが一般的である。正常値は文献によって異なるが,哺乳類の場合は 4 - 7 mM,5 - 10 mM などである。
細胞培養の培地は,糖の濃度が 1 - 4.5 g/L(5.5 - 25 mM)程度に調整されている(18)。目的によって,グルコース以外の糖を含む場合もある。
> ストレスで血糖値は増大する。コルチゾル cortisol の影響によるところが大きいとされる。
高血糖の影響: glucose toxicity
血液中に高濃度のグルコースによる
> ラットで大脳皮質の血流(cerebral blood flow, CBF)を下げる作用がある(15, 11D)。
: 文献 15 では,血糖値 8 mM にした場合,20分後に CBF が平常時の 65 %にまで低下している。
> マンニトールでも CBF が低下する。浸透圧の増大が原因と考えられている(15)。
トレース実験におけるラベリング(脳)
脳 brain では,グルコースはグルタミン,グルタミン酸,GABA,アスパラギン酸 Asp,NAA,乳酸 lactate などへ代謝される。13C 標識したグルコースを静脈に注入し,他の代謝産物(Glu, Gln, GABA, lactate etc.)に 13C が取り込まれる速度を測定することで,代謝をリアルタイムにモニターする実験系がある。
グルコース中の 13C の位置を変えることで,グルコースがどの代謝経路を通ってこれらに変換されるかを調べることが可能である。
[1-13C]Glc
Glu-C4 に入る経路(解糖 -> TCA 回路)
> 解糖によってピルビン酸の 3 位に 13C が入る。これがアセチル CoA と α-KG を経て Glu C4 に入る(1)。
: ラット,ヒトで in vivo でトレースしたデータがある。
: [1-13C]Glc infusion から数分後には [4-13C]Glu が観察され,20 分ほどで平衡に達する(4R)。
: 注入したグルコースの多くはこの経路を辿る。
> α-KG から Glu への変換は,TCA回路 の回転速度(TCA flux)に比べて十分に早いことがわかっている。
> したがって,[1-13C]Glc が減って [4-13C]Glu が増える速度が,TCA flux を示すことになる(1)。
Glu-C3 に入る経路(TCA 回路の 2 周目)
> [1-13C]Glc の一部は,以下の反応で [3-13C]Glu にも代謝される。
: α-KG の全てが Glu になるわけではなく,一部はそのまま TCA 回路を回る。
: TCA 回路では,[4-13C]α-KG は左右対称な succinate になる。13C は 2 および 3 位に入る(2)。
: 2 周目の TCA 回路では α-KG の 2 および 3 位,Glu の 2 および 3 位にも13C が入る(2)。
> 13C-MRS で調べると,ヒトでは [1-13C]Glc 注入から 60 分で [4-13C]Glu が飽和した(16)。
: [3-13C]Glu の飽和はそれよりも遅く,110-140 分後であった。
Glu-C2 に入る経路(TCA 回路の 2 周目と anaplerosis)
[1-13C]Glc が [2-13C]Glu に代謝される経路には以下の 2 つがあり(8),区別することができない。
> 第一は上に示した「C3に入る経路」で,succinate が対称なので 2 位にも入る。
: この経路は,ニューロンおよびアストロサイトの両方に存在する。
> 第二は補充反応 anaplerosis による経路である。この経路はアストロサイトにしか存在しない。
: ピルビン酸は,ピルビン酸カルボキシラーゼ PC によってオキサロ酢酸としても TCA 回路に入る。
: この場合,Glu の 2 位がラベルされることになる。
GABA-C2に入る経路(Glu-C4からGADによって合成)
> [1-13C]Glc は上記の経路で Glu C4 に入る。
> GABAergic neuron では,Glu decarboxylase (GAD) がグルタミン酸の脱炭酸によりGABAを作る。
> このとき,Glu-C4 は GABA-C2に相当する。
> また,Glu-C2 は GABA-C4 に,Glu-C3 は GABA-C3 に相当する。-> GABAの生合成を参照
[1,6-13C]Glc
> 解糖系で Glc が 2 分子に開裂する際,1 位と 6 位の C は両方とも GAP の 3 位(リン酸化 C )に入る。
> この 2 分子の GAP は化学的に同じであり,そのまま TCAサイクル に入る。
> したがって[1-13C]Glcと同様に [4-13C]Glu に代謝され,ラベルの強さは [1-13C]Glc の 2 倍になる(7)。
> 結果として SN 比が上がり,高価ではあるが良好な結果が得られる可能性が上がる。
> 上記の通り,Glu C4 のラベルを介して [2-13C]-GABA にもなる。
[2-13C]Glc
C5に入る経路(解糖->TCA回路)
> 解糖系でグルコースが2分子に開裂する際,2位と5位のCは両方ともGAPの2位に入る。
> 最終的にはピルビン酸の2位がラベルされる。
> ピルビン酸がアセチルCoAを経てTCA回路に入った場合,2位のCはα-KGおよびGluの5位に入る。
> しかし,α-KGのC5は第4段階(サクシニルCoAの生成)でCO2になって排出される。
> Gluとα-KGは平衡状態にあるので,事実上C5がラベルされたGluは蓄積しない?
C3に入る経路(補充反応 anaplerosis)
> ピルビン酸は,anaplerosis という反応でオキサロ酢酸としてもTCA回路に入る(詳細はPC)。
> このとき Glu および Gln の2位と3位がラベルされる。
> したがって,2位がラベルされたGluが蓄積する速度は,直接 anaplerosis の速度を表すことになる。
> 脳神経系では alaplerosis はアストロサイトのみに存在し,ニューロンには存在しない(1)。
: astrocyteのみで[3-13C]-Glu poolを作り,neuronには蓄積しない。
: グルタミン酸からのグルタミン合成によって,[3-13C]-Gluは容易に[3-13C]-Glnになる。
> ただし,astrocyte はグルコース取り込みが neuron よりも活発でないので,蓄積速度は遅い。
: [1-13C]-Glcは,infusion後すぐに15 - 30%に達するが,[2-13C]Glcは数パーセントである(3)。
[5-13C]Glc
> 解糖系でグルコースが2分子に開裂する際,2 位と 5 位の C は両方とも GAP の 2 位に入る。
その他考えるべきこと
> 13C-グルコースを注入することで動物が高血糖になるのは,この実験系の問題点の一つである。
> 注入速度を徐々に遅くすることで,実験の間は血糖値を 13 mM 前後に保つことができる(4R)。
> 血中グルコースの enrichment に比べて,Glu enrichment は少し低い値を示す(16)。
: Pentose shunt,非ラベルされた乳酸またはケトン体からの合成が考えられる。
References
- Sibson 2004a (Book). Chapter 5. Brain Energetics & Neuronal Activity, 2004.
- Mason 1992a. J Cereb Blood Flow Metab, 12, 434-447.
- Mason 2004a (Book). Chapter 4. Brain Energetics & Neuronal Activity, 2004.
- Fitzpatrick et al. 1990a. The flux from glucose to glutamate in the rat brain in vivo as determined by 1H-observed, 13C-edited NMR spectroscopy. J Cereb Blood Flow Metab, 10, 170-179.
- Boumezbeur et al. 2010b. Altered brain mitochondrial metabolism in healthy aging as assessed by in vivo magnetic resonance spectroscopy. J Cereb Blood Flow Metab, 30, 211-221.
- Berg et al. Biochemistry
- de Graaf 2011a (Review). State of the art direct 13C and indirect 1H-[13C] NMR spectroscopy in vivo. A practical guide. NMR Biomed 24, 958-972.
- de Graaf et al. 2003a (Review). In vivo 1H-[13C]-NMR spectrometry of cerebral metabolism. NMR Biomed 16, 339-357.
- Jitrapakdee et al. 2006a (Review). Anaplerotic roles of pyruvate carboxylase in mammalian tissues. Cell Mol Life Sci 63, 843-854.
- Govindaraju et al. 2000a. Proton NMR chemical shifts and coupling constants for brain metabolites. NMR Biomed 13, 129-153.
- Chouwdhury et al. 2008a. Chronic riluzole treatment increases glucose metabolism in rat prefrontal cortex and hippocampus. J Cereb Blood Flow Metab 28, 1892-1897.
-
Jucker et al. 1997a. 13C and 31P NME studies on the effect of increased plasma free fatty acids on
intramusclular glucose metabolism in the awake rat. J Biol Chem 272, 10464-10473.
- Tomasi et al. 2013a. Energetic cost of brain functional connectivity. PNAS early edition.
- Levin et al. 1989a. Differences in saccharin-induced cerebral glucose utilization between obesity-prone and -resistance rats. Brain Res 488, 221-232.
- Duckrow. 1995a. Decreased cerebral blood flow during acute hyperglycemia. Brain Res 703, 145-150.
- Gruetter et al. 1994a. Localized 13C NMR spectroscopy in the human brain of amino acid labeling from D-[1-13C]Glucose. J Neurochem 63, 1377-1385.
- Gurst 1991a. NMR and the structure of D-glucose. J Chem Educ 68, 1003-1004.
- Sigma-Aldrich. Pdf Link.