解糖系 Glycolysis
- 全体像
- 酵素の一覧
- 代謝産物の流入および流出
- 各反応の解説
- 覚え方(もしもし解糖系)
全体像
解糖系とは,グルコース glucose が2分子の ATP を生み出しつつ各種酵素で分解され,2分子のピルビン酸 pyruvate を生じる一連の反応である(1)。生化学的には,2分子の NADH(酸化的リン酸化などに使われる)が生じることも重要である。
"Glycolysis is the sequence of reactions that metabolized one molecule of glucose to wto molecules of pyruvate with the concomitant production of two molecules of ATP" (1).
- この反応が酸素を必要としないのは,大気中の酸素濃度が増える前に生まれた経路だからと考えられる(1)。→ See 解糖系の進化
- 解糖系を逆に辿ると糖新生経路に近いが,いくつかの高エネルギー反応の部分で迂回経路を通っている(1)。
- 解糖と糖新生は,同じ細胞内で同時には起こらない(1)。
-
解糖系の中間代謝産物の濃度は,一般に非常に低い(0.1 mmol/kg 以下)。つまり各反応が早い(3)。

Molecular Biology of the Cell, 5th ed.
全体の反応 net reaction は
Glucose + 2 Pi + 2 ADP + 2 NAD -> 2 pyruvate + 2 ATP + 2 NADH + 2 H+ + 2 H2O
である(1)。
最終産物であるピルビン酸 pyruvate は,原則として以下のように代謝される(1; 図は 7)。
- 好気的条件下では,アセチル CoA を経てミトコンドリアの TCA 回路に入る。
- 嫌気的条件下では,細胞質で乳酸 lactate に変換される(乳酸発酵)。
- 酵母をはじめとしたいくつかの生物では,細胞質でエタノール ethanol に変換される(アルコール発酵)。

酵素の一覧
- ヘキソキナーゼ Hexokinase, HX
- ホスホグルコースイソメラーゼ Phosphoglucoseisomerase
- ホスホフルクトキナーゼ Phosphofructokinase, PFK
- アルドラーゼ Aldolase
- トリオースリン酸イソメラーゼ Triose phosphate isomerase
- グリセルアルデヒド3-リン酸デヒドロゲナーゼ Glyceraldehyde 3-phosphate dehydrogenase, GAPDH, G3PDH
- ホスホグリセリン酸キナーゼ Phosphoglycerate kinase, PGK
- ホスホグリセリン酸ムターゼ Phosphoglycerate mutase, PGM
- エノラーゼ Enolase
- ピルビン酸キナーゼ Pyruvate kinase, PK
解糖系への代謝産物の流入および解糖系からの流出
流入
| 他の糖 | 説明 |
|
フルクトース Fructose |
肝臓とその他の組織で,異なる経路で解糖系に入る(1)。
|
|
ガラクトース Galactose |
ガラクトース-1-リン酸 > グルコース-1-リン酸 > グルコース-6-リン酸 と変化して解糖系に入る(1)。 |
|
マンノース Mannose |
Hexokinase が マンノース-6-リン酸にリン酸化し,これが phosphomannnose isomerase によって G6P になり解糖系に入る。 |
|
DHAP |
脂肪分解 lipolysis で生じたグリセロールは,glycerol kinase および glycerol phosphate dehydrogenase によって DHAP となる。ここから解糖によってピルビン酸になる場合と,糖新生によってグルコースになる場合がある。グリセロール合成の場合も,DHAP から分岐する。 |
流出
| 代謝産物 | 説明 |
|
G6P |
グリコーゲン合成: G6P のリン酸基がホスホグルコムターゼ phosphoglucomutase によって 1 位に転移すると G1P ができる。G1P は UDP との反応を経てグリコーゲン鎖に付加される。 |
|
G6P |
ペントースリン酸回路: G6P が G6PD によって脱水素されると,NADP+ が NADPH に還元されるとともに,6-phosphoglucono-δ-lactone ができる。炭素骨格がペントースリン酸回路へ流れる経路である。 |
|
DHAP |
脂肪合成: 主要な貯蔵資質であるトリアシルグリセロール TG は,グリセロールと脂肪酸がエステル結合した分子である。 DHAP はグリセロール合成の原料となり,最終的に TG に取り込まれる。 |
各反応の解説
1. 解糖の開始
トランスポーター GLUT によって細胞内に取り込まれたグルコースは,ヘキソキナーゼ hexokinase によって 6 位でリン酸化され,G6P となる。このリン酸基の付加によってグルコースの安定性が低下し,その後の代謝反応を受けやすくなる。
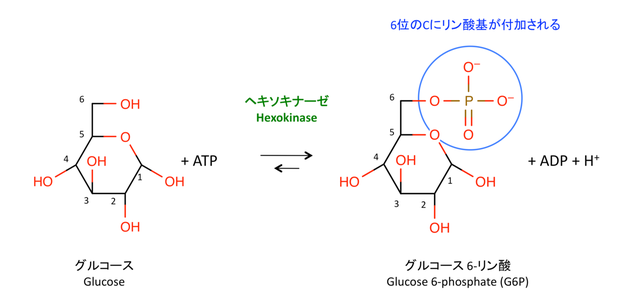
> Hexokinase は,他の多くのキナーゼと同様に,反応に Mg2+ を必要とする。
: G6P はヘキソキナーゼをアロステリック阻害する(4I)。
: Hexokinase 阻害によって解糖系の flux を低下させると,一般に寿命は長くなる。
> この反応は不可逆的で,hexokinase は 以下の PFK, PK とともに 3 つの解糖系の律速酵素の一つである(1)。
: 詳細は 解糖系の活性調節 を参照のこと。
> G6P はグリコーゲン合成の出発点になる。
: G6P のリン酸基がホスホグルコムターゼによって1位に転移して G1P ができる。
: G1P は UDP との反応を経てグリコーゲン鎖に付加される。詳細はグリコーゲンを参照のこと。
: 糖尿病患者の筋肉で glycogen が少ないが,これは合成ではなく Glc のり込み低下が原因(2)。
> G6P は ペントースリン酸回路の出発点になる(1)。
: ペントースリン酸回路は,補酵素 NADPH と,核酸などの原料になる ribose を産生するための経路。
: NADPH が必要な場合 (= NADP+ が多い),G6PD が活性化しペントースリン酸経路へ多量の G6P が流れる。
2. フルクトースへの変換: Isomerization
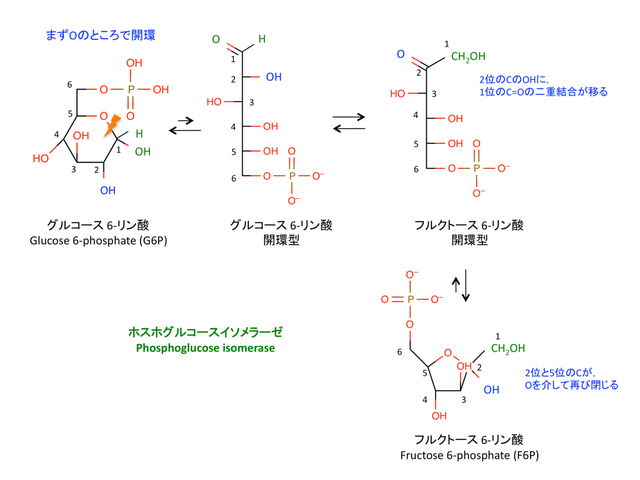
> これは isomerizationという反応で,phosphoglucose isomerase によって触媒される。
> G6P はアルデヒド基をもつ糖であるアルドース aldose,F6P はケトース ketoseである。
3. フルクトース-6-リン酸のさらなるリン酸化
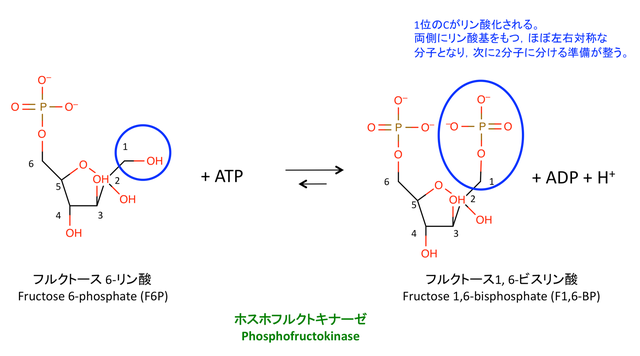
> Phosphofrukutokinase(PFK)によるこの反応は,解糖系の 律速段階 として様々な分子による調節を受ける。
: たとえば cAMP, ADP により活性化され,クエン酸 citrate, ATP, プロトン, 長鎖脂肪酸により抑制される。
: 詳細は PFK のページ および 解糖系の活性調節 のページを参照のこと。
: このほか,hexokinase および pyruvate kinase による反応が不可逆である。
> 摂取されたフルクトースは,肝臓以外の組織では hexokinase によってリン酸化され,解糖系に合流する(1)。
: 1 位 -> 6 位の順にリン酸化され,F1,6-BP として解糖系に入ってくる。
4. フルクトース1,6-ビスリン酸の開裂
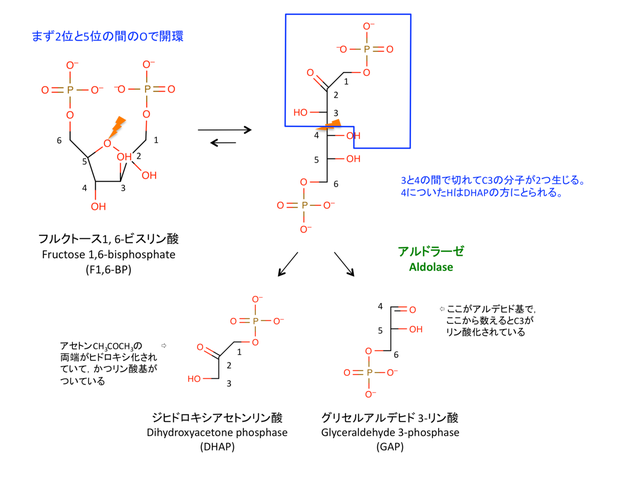
> ここまでの反応で,グルコース 1 分子あたり 2 分子の ATP を消費していることになる。
> アルドラーゼに触媒される開裂反応は,可逆的である。
> GAP はそのまま分解が進むが,DHAP は次の反応で GAP に変換される。
> DHAP はグリセロール-3-リン酸を経てグリセロールになり,トリアシルグリセロール TAG の合成に寄与する。
: この反応については,グリセロールのページを参照のこと。
5. DHAPの異性化
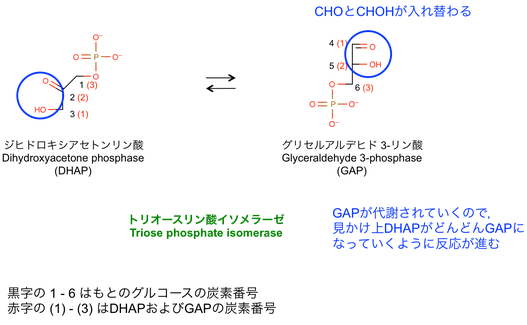
> DHAP はトリオースリン酸イソメラーゼ triose phosphase isomerase によって GAP に変換される。
: GAP と DHAP の変換は,可逆的で非常に早い反応である。
: 平衡は DHAP に偏っているが,GAP は反応 6 で取り除かれるので結果的に全体として DHAP が GAP に変わっていく。
ここで同一の分子を作り出せるように,これまでの反応(フルクトースへの変換など)が進められてきたと考えてみてもよい。
> 肝臓では,フルクトースは fructokinase は開裂後に DHAP として解糖系に合流する(1)。
: Fructokinase によって F1P にリン酸化され,glyceraldehyde および DHAP に開裂する。
: DHAP はステップ 5 に合流する。
: Glyceraldehyde は triose kinase によって 3 位でリン酸化され,GAP として解糖系に入る。
6. GAPのリン酸化
この反応は 2 段階である。
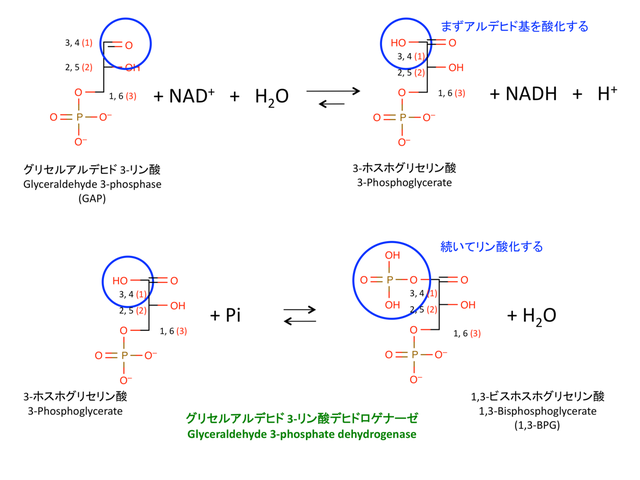
> 高いリン酸基転移ポテンシャルをもつ 1,3-ビスホスホググリセリン酸が生じる。これは ADP に Pi を付加できる。
: これはつまり,GAPDH によるリン酸化が分子にかなりのエネルギーを与えているということになる。
: 実際,2 番目の反応はかなりの uphill reaction である。
: これには,GAPDH が高エネルギーのチオエステル結合を含む中間体になることによって成される。
: チオエステル結合のもつ高エネルギーについては,化学結合のページを参照のこと。
> Glycerate 3-phosphate も高エネルギー中間体である。
> この反応を触媒する酵素 GAPDH (G3PDHとも) には,少なくとも 5 つの分子種がある。
: 上記の反応は,EC.1.2.1.12 G3PDH (phosphorylating) によるものである。
> 解糖系で唯一 NADH を生じる重要なステップだが,NAD+が常に供給されないとこの反応は進まない。
: NADH を酸化して NAD+ にする経路: アルコール発酵,乳酸発酵。
> Pi はどこから供給されるのか?
7. 1,3-bisphosphoglycerateからADPへのリン酸基の転移
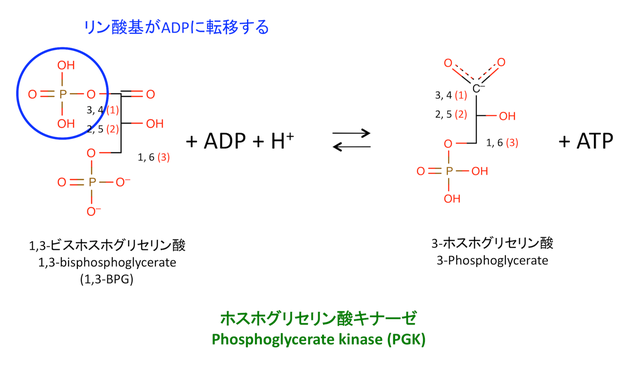
> 1 分子の ATP が生じる。前の反応でつけたリン酸基を ADP に移しているだけ。
: これは 1,3-BPG が,数少ない「ATP よりもエネルギー準位の高い分子である」から起こりうる反応。
: 解糖系の中間体で ATP よりもエネルギー準位が高いのは,この分子と PEP のみ。
: ATP よりもエネルギー順位が高い中間体が 2 個ある = 「解糖系では 2 分子の ATP が作られる」
> 反応には Mg2+ が必要。
8. リン酸基の分子内転移

> PGM はリン酸化 Ser をもっており,これでまず基質を二リン酸にする(1)。次に余分なリン酸基を回収する。
> ムターゼ mutase とは,分子内で官能基を移動させる酵素の総称である。英語では [mjuːteiz] と発音する。
> PGM はグリコーゲン分解で登場するホスホグルコムターゼ phosphoglucomutase とは違うので注意(1)。
9. 脱水反応
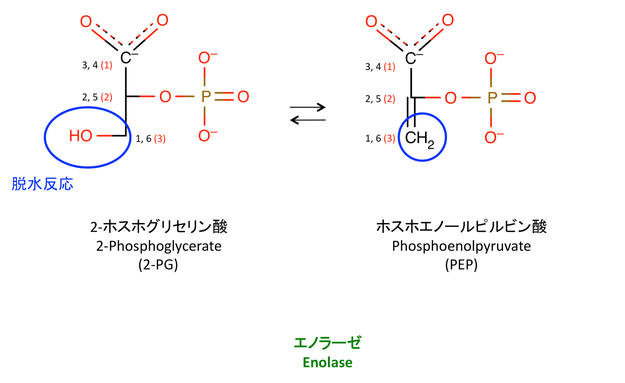
右辺の + H2O は省略(書き忘れ)。シンプルな脱水反応である。
> エノール enol は,アルケンの二重結合の片側にヒドロキシ基が置換した アルコール のこと(1)。
: PEP はエノール型のピルビン酸にリン酸基(phospho)がついたもの。
: 通常は,酸素の方が電気陰性度が高いので,エノールの二重結合が O の方へ引っ張られケト型になる。
: リン酸基があることで不安定な enol 型が保たれ,PEP として存在できる。
: 次の反応でリン酸基が取られると,ピルビン酸はケト型へと変化する。
10. PEP から ADP へのリン酸基の転移
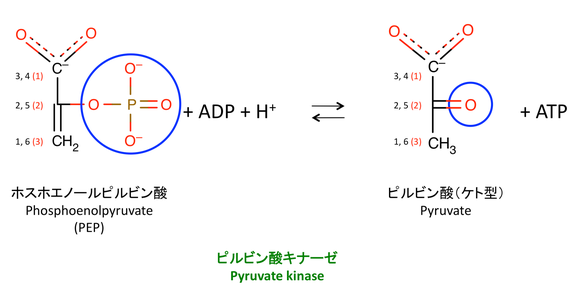
ホスホエノールピルビン酸から ADP にリン酸基を転移させることで,ATP を作る。この反応はピルビン酸キナーゼ pyruvate kinase によって触媒される。
- ピルビン酸 pyruvate は,好気的条件下ではピルビン酸デヒドロゲナーゼ PDH によってアセチル CoA になり,ミトコンドリアにある TCA回路 に入る。
- ピルビン酸は,monocarboxylate transporters (MCTs), とくに MCT1 によってミトコンドリアまで輸送される(5I)。
- 嫌気的条件化では,乳酸デヒドロゲナーゼ LDH によって乳酸 lactate になる。
> この反応は不可逆であり,PK は hexokinase および PFK とともに 3 つの解糖系の律速酵素の一つである(1)。
: 詳細は 解糖系の活性調節 を参照のこと。
> 直接は関係ないが,PEP は C4 植物で葉緑体へ二酸化炭素を運ぶためのキャリアとしても使われる(1)。
覚え方
もしもし解糖系(知識の泉 さん)
もしもしかめよ〜の音程で歌いましょう。
1.
グルグル6リン フル6リン フルクの1, 6ビスリン酸
アルドで2つに脱離して -2のATP
2.
次に1, 3ビスホスホ +2のATP
3ホス 2ホスエノラーゼ 血糖値はココ阻害
3.
ホスホのエノールピルビン酸 ココでは後に戻れない
+2のATP 作ってなるのがピルビン酸
References
- Berg et al. Biochemistry: 使っているのは 6 版ですが 7 版を紹介しています。
- Shulman 1995a (Review). In vivo regulation of muscle glycogen synthase and the control of glycogen synthesis. Proc Natl Acad Sci USA, 92, 8535-8542.
- de Graaf et al. 2003a (Review). In vivo 1H-[13C]-NMR spectrometry of cerebral metabolism. NMR Biomed 16, 339-357.
- Jucker et al. 1997a. 13C and 31P NME studies on the effect of increased plasma free fatty acids on intramusclular glucose metabolism in the awake rat. J Biol Chem 272, 10464-10473.
- Ling et al. 2012a. D-Lactate altered mitochondrial energy production in rat brain and heart but not liver. Nutr Metab 9, 6.
- Wei et al. 2009a. Tor1/Sch9-regulated carbon source substitution is as effective as calorie restriction in life span extension. PLoS Genet 5, e1000467.
- "TumorMetabolome" by Kathleen A Vermeersch, Mark P Styczynski - DOI: 10.4103/1477-3163.113622; PMID: 23858297. Licensed under CC BY-SA 3.0 via Commons.