アセチルCoAカルボキシラーゼ,ACC
概要
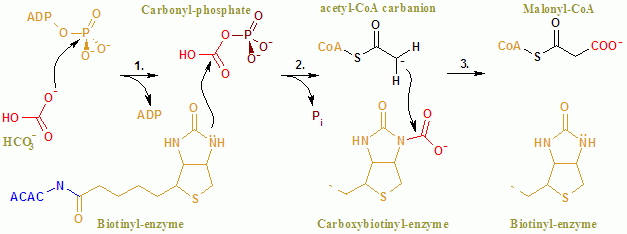
アセチル CoA カルボキシラーゼ(EC 6.4.1.2)は,アセチル CoA からマロニル CoA を作る酵素で,脂肪酸合成の律速酵素 であるとともに,β 酸化にも関わっている。 触媒する反応は,「カルボキシラーゼ」 の名前の通りアセチル CoA に CO2 を付加する反応であり,以下のようなステップで行われる(4,5)。
- 青字の ACAC が酵素で,ビオチン biotin 化されている。
- ATP のリン酸基が炭酸水素イオン bicarbonate に移り,これがビオチン部分と結合する。反応性の高い CO2- が生じる。
- これがアセチル CoA を攻撃し,CO2- が転移することでマロニル CoA になる。
この反応は,ATP および bicarbonate 依存的である。
> マロニル CoA は CPT を阻害し,β 酸化を抑制する。
: つまり,脂肪酸合成が起こっているときには,酸化は起こらないようになっている。
: 合成は細胞質,酸化はミトコンドリアだが,同じ細胞内で逆の反応が起こらないという基本を守っている。
> マロニル CoA による β 酸化の阻害は,筋肉でも起こる(6)。
: 筋肉では,脂肪酸合成はほとんど起こらないと考えられている。
: 筋肉で発現している ACC は,β 酸化の制御因子としての機能を果たしていると考えられる。
ACC の活性制御機構
アロステリック制御
> クエン酸 citrate のような tricarbocxylic acid で活性化される(5)。
: 実験的に活性を測定する場合には,通常 citrate を反応液に加える。
: Citrate が多いということは,TCA 回路が活発である,つまりエネルギーが余っている状態である。
> 長鎖アシル CoA チオエステルで阻害される(6)。
: Palmitoyl CoA は ACC を阻害するとともに,ミトコンドリアからの citrate 輸送を阻害する。
: Citrate は細胞質でアセチル CoA に変換され,ACC の基質となる。
: Palmitoyl CoA はさらに NADPH 合成を阻害する。ペントースリン酸回路の G6PDH を阻害。
: まとめると,酵素を不活性化し,さらに原料を絶つという強力な阻害である。
ホルモンによる制御
> AMPK にリン酸化されると不活性化する(6)。脱リン酸化で活性型になる。
: AMPK は,エネルギー不足のときに AMP で活性化される。
: よってエネルギーが足りないときに ACC は不活化され,脂肪は作られない。
> 成長ホルモン GH は,ブタの脂肪組織で ACC の活性,mRNA量,タンパク質量を低下させる(3)。
> エピネフリン,グルカゴンは ACC を不活性化する(6)。
: これらは脂質を動員,利用を促進するホルモンなので,合成を阻害するのは当然である。
> インスリンは ACC を脱リン酸化して活性化する(6)。
ノックアウトマウス Knockout mice
ACC1 KO mice
> ACC1-/- miceはembryonic lethalである(2)。
> ACC1+/- miceは,ACC1のmRNA量は半分だがリン酸化ACC量は変わらず,補償が働いている(2)。
: 肝細胞へのアセチルCoAの取り込み量,マロニルCoA量,体重がwtと変わらなかった。
ACC2 KO mice
> 絶食によって,肝臓,筋肉,心臓のmalonyl-CoA量が低下する(1)。
> ACC2 KOは,肝臓でのマロニルCoA量は変わらないが,心臓と筋肉では低下(1)。
> マロニルCoAはミトコンへの脂肪酸輸送を阻害するので,KOではβ酸化が盛んだろう(1)。
> ACC2 KO miceの肝臓は脂質含量が低い。ただしACC2は肝臓にはなく二次的効果(1)。
> ACC2 KOは,wtよりも摂食量が多いが体重は軽い。脂肪組織量が少ない(1)。
その他未整理
> 肝臓,脂肪組織で発現する細胞質型の ACC1 と,心臓,筋肉のミトコンに局在する ACC2 がある(1)。
> マロニル CoA には ACC1 が作る細胞質プールと,ACC2 が作るミトコンプールがある(2)。
References
- Abu-Elheiga 2001a. Science, 291, 2613-2616.
- Abu-Elheiga 2005a. Mutant mice lacking acetyl-CoA carboxylase 1 are embryonically lethal. PNAS, 102, 12011-12016.
- Chaves et al. 2013a (Review). The metabolic effects of growth hormone in adipose tissue. Endocrine 44, 293-302.
- "ACAC mechanism" by Original uploader was BorisTM at en.wikipedia - Transferred from en.wikipedia; transfer was stated to be made by User:xvazquez.. Licensed under Public Domain via Wikimedia Commons.
- Volpe and Vagelos 1976a (Review). Mechanism and regulation of biosynthesis of saturated fatty acids. Physiol. Rev. 46, 339-417.
- Berg et al. Biochemistry: 使っているのは 6 版ですが 7 版を紹介しています。