AMP-activated protein kinase (AMPK),哺乳類
概要
細胞のエネルギーセンサーとして重要なセリン/スレオニンキナーゼ。シグナル伝達による制御のほか,AMP/ATP比が上がる(エネルギーとしてのATPが消費される)ことで活性化する。
生理機能は多岐にわたるが,基本的には糖や脂質を酸化し,ATPを生み出す方向に作用する。また,視床下部を刺激して食欲を増進する作用もある(3I)。
酵母から哺乳類まで存在する保存性の高い分子である(1)。
構造
> α(活性サブユニット),β,γ(両方とも調節サブユニット)から成る三量体タンパク質(3I)。
> αサブユニットは,N末端にキナーゼドメインがあり,C末端でβ,γサブユニットと複合体を形成(1)。
> α1, α2, β1, β2, γ1, γ2, γ3のアイソフォームが存在する(1)。
シグナル伝達による活性調節
AMPKをリン酸化するキナーゼ: LKB1,CaMKK α/β、TAK1 (TGF-β-activated kinase) が知られている(1)。
LKB1
> LKB1は、Ser431がERKにリン酸化されることで核から細胞質へ移行し,MO25 (mouse protein 25) およびSTRAD (STE-20-related kinase adapter alpha)と結合してAMPKをリン酸化する(1)。
AMPによるアロステリック調節
> γサブユニットには4個のCBS (cystathionine-beta-synthase) ドメインがあり,ATPまたはAMPと結合(1)。
> AMPと結合している場合に活性化する。空腹,運動などでAMP/ATP比が上がった場合に活性化する(1)。
肝臓の脂質代謝における役割
AMPKは細胞のエネルギー状態が低いときに活性化する。足りないエネルギーを補充するために,肝臓では脂肪酸合成を抑制し,かつ脂肪酸の酸化を促進する。
脂肪酸合成の抑制
AMPKの活性が高い状態では,脂肪酸合成経路への原料の流入が阻害される(1)。これにはいくつかのメカニズムがある。右図(文献2, Abu-Elhaiga 2005a)のように,脂肪酸はアセチルCoAからACC,FASなどの作用によって合成されるが,AMPKは
- アセチルCoAからの脂肪酸合成の律速酵素,ACC をリン酸化して活性を低下させる。
- ACCによって合成されるマロニルCoAを脂肪酸合成に乗せないように,マロニルCoAカルボキシラーゼの発現を増大させる。
- 脂肪酸合成酵素 FAS の発現を低下させる。
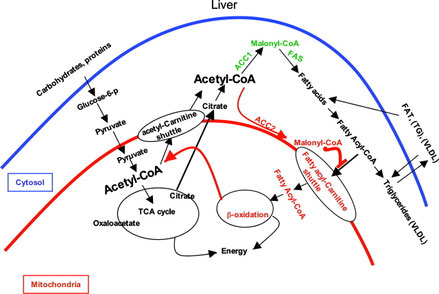
脂肪酸酸化の促進
> ACCによってアセチルCoAから作られるマロニルCoAは,それ自身が脂肪酸酸化を抑制する作用をもつ(1)。
: 脂肪酸は,CoA と結合してfatty-acyl CoAになったあと,ミトコンドリアに運ばれて酸化される。
: ミトコンドリアへの運搬は,CPT1が行うが,マロニルCoAはその活性を抑制する作用がある。
: したがって,AMPKがACCをリン酸化することでマロニルCoA量が減ると,結果的に脂質酸化もupする。
References
- 浅野知一朗 2009a (Review). AMPK を介するエネルギー代謝調節. 実験医学 増刊 27 73-77. 肥満・糖尿病の病態を解明するエネルギー代謝の最前線.
- Abu-Elheiga 2005a. Mutant mice lacking acetyl-CoA carboxylase 1 are embryonically lethal. PNAS, 102, 12011-12016.
- Apfeld et al. 2004a. The AMP-activated protein kinase AAK-2 links energy levels and insulin-like signals to lifespan in C. elegans. Genes Dev 18, 3004-3009.